Cytokinesis has to be closely coordinated with chromosome replication and segregation to generate viable offspring that contains the full complement of hereditary information. In most bacteria, this process is mediated by a ring-shaped multi-protein complex, called the divisome, which spans all layers of the cell envelope. The basis of the divisome is formed by polymers of the tubulin homologue FtsZ that, directly or indirectly, recruit all other divisome components and, furthermore, play a pivotal active role in the subsequent constriction process. Given its fundamental role in cell division, FtsZ is the primary target of systems involved in the temporal and spatial regulation of cytokinesis.
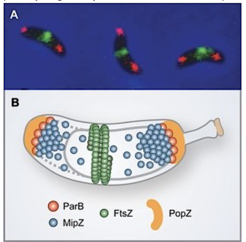
Division site placement by MipZ. (A) In vivo localization of FtsZ (green) and ParB (red). (B) Schematic showing the components of MipZ system.
Division site placement by MipZ. (A) In vivo localization of FtsZ (green) and ParB (red). (B) Schematic showing the components of MipZ system.
We have previously identified a novel regulatory mechanism that controls the positioning of the division plane in C. crescentus and probably many other alpha-proteobacterial species (Thanbichler & Shapiro, 2006). It is founded on the P-loop ATPase MipZ, which forms a dynamic complex with the DNA partitioning protein ParB at the chromosomal origin of replication (Figure 2). In the predivisional cell, the two copies of the origin/ParB complex are sequestered to opposite cell poles through interaction with the scaffolding protein PopZ. As a consequence, MipZ forms a gradient within the cytoplasm, with its concentration being highest at the tips of the cell and lowest at the cell center. MipZ acts as a direct inhibitor of FtsZ polymerization, thus limiting formation of the division apparatus to the midcell region.
Steady-state protein gradients are a well-known regulatory strategy in eukaryotic cells, usually established by localized protein synthesis followed by subsequent diffusion and protein degradation. However, such diffusional gradients are extremely unstable at the scale of a small prokaryotic cell, suggesting that formation of the MipZ gradient is based on a different principle. We have solved the crystal structures of MipZ in the apo and ATP-bound form and analyzed the mechanism of MipZ localization (Kiekebusch et al., 2012). Our studies showed that MipZ serves as a molecular switch that, similar to small GTPases, uses nucleotide binding and hydrolysis to alternate between two different functional states with distinct interaction networks and diffusion rates. This behavior drives a unique localization cycle, with MipZ molecules oscillating back and forth between the polar ParB complexes and pole-distal regions of the nucleoid. These findings for the first time shed light on the basis of steady-state gradient formation in bacteria and might provide a general mechanistic framework for other gradient-forming systems in both prokaryotic and eukaroytic cells (Kiekebusch and Thanbichler, 2013).
Currently, we are aiming at a better understanding of the mechanistic basis of MipZ gradient formation, in particular by studying the regulatory interplay between MipZ and its various cellular interaction partners.
 In vivo localization of FtsZ (green) and ParB (red). (B) Schematic showing the components of MipZ system.")